Recommendations for nomenclature and tables in biochemical thermodynamics
(Recommendation 1994)
New Recommendations 4.1
Continued from 3 Corresponding New Recommendations
Contents of Section
4 Additional New Recommendations
4.1 Recommendations concerning chemical reactions
References for this Section
4. Additional New Recommendations
4.1 Recommendations Concerning Chemical Reactions
The thermodynamics of reactions of species in aqueous solution is discussed in every textbook on physical chemistry, but this section is included to contrast the nomenclature with that of the next section and to respond to the special needs of biochemistry. As mentioned in Section 3, equilibrium constants of chemical reactions that are used in biochemistry are taken to be functions of T, P, and I. Therefore, the standard thermodynamic properties are also functions of T, P, and I. The standard Gibbs energy of reaction Dr G o for reaction 9 is calculated using
Dr G o = - RT ln K . . . . . . . . (11)
and there are corresponding values of Dr H o and Dr S o that are related by
Dr G o = Dr H o - TDr S o . . . . . . . . (12)
The standard enthalpy of reaction is given by
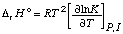 . . . . . . . . (13)
. . . . . . . . (13)
If Dr H o is independent of temperature in the range considered, it can be calculated using
Dr H o = [RT1T2/(T2 - T1)] ln (K2/K1) . . . . . . . . (14)
If the standard molar heat capacity change Dr CP o is not equal to zero and is independent of temperature, the standard molar enthalpy of reaction varies with temperature according to
Dr Ho(T) = Dr Ho(T *) + Dr CPo(T - T *) . . . . . . . . (15)
The reference temperature T * is usually taken as 298.15 K. In this case, Dr G o and K vary with temperature according to (ref. 6)
Dr G o(T) = - RT ln K(T )
= Dr Ho(T *) + Dr CPo(T - T *) + T {Dr G o(T *) - Dr H o(T *)}/T * - T Dr CPo ln (T/T *) . . . . . . . . (16)
Additional terms containing (![[del]](thgif/del.gif) Dr CPo/T )P and higher order derivatives may be needed for extremely accurate data or for a very wide temperature range.
Dr CPo/T )P and higher order derivatives may be needed for extremely accurate data or for a very wide temperature range.
Equations 13 and 14 are exact only when the equilibrium constants are based on a molality standard state. If the equilibrium constants were determined with a standard state based on molarity, these equilibrium constants should be converted to a molality basis prior to using equation 13. For dilute aqueous solutions, mi = ci/r where mi and ci are, respectively, the molality and the molarity of substance i and r is the mass density of water in kg L-1. If this conversion is not made, there is an error of RT 2(lnr /T)P,I for each unsymmetrical term in the equilibrium constant. This quantity is equal to 0.187 kJ mol-1 for dilute aqueous solutions at 298.15 K. Similar statements pertain to equations 27 and 28, which are given later in this document.
Since the standard thermodynamic properties Dr G o and Dr H o apply to the change from the initial state with the separated reactants at c o to the final state with separated products at c o, it is of interest to calculate the changes in the thermodynamic properties under conditions where the reactants and products have specified concentrations other than c o. The change in Gibbs energy Dr G in an isothermal reaction in which the reactants and products are not all in their standard states, that is, not all at 1 M, is given by
Dr G = Dr Go + RTlnQ . . . . . . . . (17)
where Q is the reaction quotient of specified concentrations of species. The reaction quotient has the same form as the equilibrium constant expression, but the concentrations are arbitrary, rather than being equilibrium concentrations. Ideal solutions are assumed. The change in Gibbs energy Dr G in an isothermal reaction is related to the change in enthalpy Dr H and change in entropy Dr S by
Dr G = Dr H - TDr S . . . . . . . . (18)
The corresponding changes in entropy and enthalpy are given by
Dr S = Dr So - RlnQ . . . . . . . . (19)
Dr H = Dr Ho . . . . . . . . (20)
The standard reaction entropy can be calculated from the standard molar entropies of the reacting species: Dr S o = S nio(i ), where ni is the stoichiometric number (positive for products and negative for reactants) of species i.
The electromotive force E of an electrochemical cell is proportional to the Dr G for the cell reaction.
Dr G = - |ne|FE . . . . . . . . (21)
where |ne| is the number of electrons transferred in the cell reaction and F is the Faraday constant (96 485.31 C mol-1). Substituting equation 17 yields
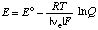 . . . . . . . . (22)
. . . . . . . . (22)
where E o = - Dr G o/|ne|F is the standard electromotive force, that is the electromotive force when all of the species are in their standard states, but at the ionic strength specified for Dr G o. The electromotive force for a cell is equal to the difference in the electromotive forces of the half cells.
The standard Gibbs energy and enthalpy of reaction can be calculated from the formation properties of the species.
Dr G o = S niDf G o(i) . . . . . . . . (23)
Dr H o = S niDf H o(i) . . . . . . . . (24)
where the ni is the stoichiometric numbers of species i. The standard entropy of formation of species i can be calculated using
Df S o(i) = Df H o(i) - Df G o(i)]/T . . . . . . . . (25)
Two special needs of biochemistry are illustrated by considering the seven species in Table I. The first part of Table I gives the standard thermodynamic properties as they are found in the standard thermodynamic tables (see Appendix). The standard thermodynamic tables give the standard formation properties for the standard state, which is the state in a hypothetical ideal solution with a concentration of 1 M but the properties of an infinitely dilute solution and the activity of the solvent equal to unity. This means that the tabulated thermodynamic properties apply at I = 0. Since many biochemical reactions are studied at about I = 0.25 M, the tabulated values of Df G o(i) and Df H o(i) in The NBS Tables of Chemical Thermodynamic Properties and the CODATA Key Values for Thermodynamics have to be corrected to ionic strength 0.25 M, as described in Section 5.3. The values at I = 0.25 M are given in the second part of Table I. No adjustments are made for H2O, MgHPO4, and glucose because the ionic strength adjustment is negligible for neutral species. We will see in Section 5.1 that the transformed Gibbs energy G ' is the criterion of equilibrium at specified pH and pMg. In Section 5.4, we will see that the calculation of transformed thermodynamic properties involves the adjustment of the standard formation properties of species to the desired pH and pMg by use of formation reactions involving H+ and Mg2+. The standard transformed formation properties of species can be calculated at any given pH and pMg in the range for which the acid dissociation constants and magnesium complex dissociation constants are known. However, for the purpose of making tables, it is necessary to choose a pH and pMg that is of general interest. For the tables given here, pH = 7 and pMg = 3 are used because they are close to the values in many living cells. Table II shows the result of these calculations for the species in Table I. The ions H+ and Mg2+ do not appear in this table because it applies at pH = 7 and pMg = 3. These methods have been applied to calculate the transformed formation properties of Pi (ref. 4); glucose 6-phosphate (ref. 5); adenosine, AMP, ADP, and ATP (ref. 7).
Table I. Standard Formation Properties of Aqueous Species at 298.15 K.
| Df Ho/kJ mol-1 | | Df Go/kJ mol-1 |
|---|
| | I = 0 M | |
| H2O | -285.83 | | -237.19 |
| H+ | 0.00 | | 0.00 |
| Mg2+ | -467.00 | | -455.30 |
| HPO42- | -1299.00 | | -1096.10 |
| H2PO4- | -1302.60 | | -1137.30 |
| MgHPO4 | -1753.80 | | -1566.87 |
| Glucose | -1262.19 | | -915.90 |
| | I = 0.25 M | |
| H2O | -285.83 | | -237.19 |
| H+ | 0.41 | | -0.81 |
| Mg2+ | -465.36 | | -458.54 |
| HPO42- | -1297.36 | | -1099.34 |
| H2PO4- | -1302.19 | | -1138.11 |
| MgHPO4 | -1753.80 | | -1566.87 |
| Glucose | -1262.19 | | -915.90 |
Table II. Standard Transformed Formation Properties of Species at 298.15 K, pH = 7, pMg = 3, and I = 0.25 M.
| Df H 'o/kJ mol-1 | | Df G 'o/kJ mol-1 |
|---|
| H2O | -286.65 | | -155.66 |
| HPO42- | -1297.77 | | -1058.57 |
| H2PO4- | -1303.01 | | -1056.58 |
| MgHPO4 | -1288.85 | | -1050.44 |
| Glucose | -1267.11 | | -426.70 |
Tables I and II can be extended by use of measured equilibrium constants and enthalpies of reaction for enzyme-catalyzed reactions. For example, the species of glucose 6-phosphate can be added to these two tables because the equilibrium constant for the hydrolysis of glucose 6-phosphate has been measured at several temperatures, and because the acid dissociation constant and magnesium complex dissociation constant for glucose 6-phosphate are known at more than one temperature. However, as the standard thermodynamic properties are not known for any species of adenosine, AMP, ADP, or ATP, it is necessary to adopt the convention that Df G o = Df H o = 0 for adenosine in dilute aqueous solution at each temperature. This convention was introduced for H+ a long time ago. This method has been used to calculate the standard enthalpies and standard Gibbs energies of formation of adenosine phosphate species relative to H2ADP- at 298.15 K (ref. 8). When this convention is used, it is not possible to calculate the enthalpy of combustion of adenosine, but it is possible to calculate Dr G o and Dr H o for reactions of adenosine that do not reduce it to CO2, H2O, and N2. If the standard thermodynamic properties of all of the species of a reactant are known, Dr G 'o and Dr H 'o can be calculated at any specified pH and pMg, as described in the next section. When Df G o and Df H o are eventually determined for adenosine in dilute aqueous solution, the values of Df G o and Df H o of the other species in the ATP series can be calculated, but this will not alter the equilibrium constants and enthalpies of reaction that can be calculated using the tables calculated using the assumption that Df G o = Df H o = 0 for adenosine.
In making these calculations, the pH has been defined by pH = - log10([H+]/c o), rather than in terms of the activity, as it is in more precise measurements. The reason for doing this is that approximations are involved in the interpretation of equilibrium experiments on biochemical reactions at the electrolyte concentrations of living cells. Even Na+ and K+ ions are bound weakly by highly charged species of biochemical reactants, like ATP. As an approximation the acid dissociation constants and magnesium complex dissociation constants are taken to be functions of the ionic strength and the different effects of Na+ and K+ are ignored. These approximations can be avoided in more precise work, but only at the cost of a large increase in the number of parameters and the amount of experimental work required.
References for this section
4. Alberty, R. A. (1992) Biophys. Chem. 42, 117-131.
5. Alberty, R. A. (1992) Biophys. Chem. 43, 239-254.
6. Clarke, E. C. W., and D. N. Glew, D. N. (1966) Trans. Faraday Soc. 62, 539-547.
7. Alberty, R. A., and Goldberg, R. N. (1992) Biochemistry 31, 10610-10615.
8. Wilhoit, R. C. (1969) Thermodynamic Properties of Biochemical Substances, in Biochemical Microcalorimetry, H. D. Brown, ed., Academic Press, New York.
Continue with 4.2 Recommendations concerning biochemical reactions.
Return to home page for Biochemical Thermodynamics.